Diferencias contra las plantas C3 y C4
Para la
transformación de hidratos de carbono usan todas las plantas verdes la
fotosíntesis. En la "fase oscura" de ésta, el dióxido de carbono se
fija y se transforma en hidratos de carbono. La mayoría de las plantas (C3)
utilizan el metabolismo C3, donde el dióxido de carbono pasa pasivamente por
los estomas de las células y durante el día se fija como sustrato en el ciclo
de Calvin.
Una adaptación de
este mecanismo se encuentra en las plantas C4, que aumentan activamente la
concentración de dióxido de carbono, con consumo energético para su fijación.
Aquí existe una separación física (dos tipos de células, células mesófilas y
las células de la vaina) para la previa fijación y metabolización del dióxido
de carbono. Esto permite a las plantas, cerrar sus estomas parcialmente, que al
contrario de las plantas C3 no se ven limitadas por la simple difusión de
dióxido de carbono en las células. Cuando los estomas están cerrados
parcialmente se reduce la pérdida de agua en la planta. Por ello se les
encuentra en zonas más secas y calientes. La fijación de dióxido de carbono en
el ciclo de Calvin es igual al de las plantas C3.
Para sobrevivir en
regiones áridas, utilizan las plantas CAM otro mecanismo, los pasos para la
fijación del CO2 del ciclo de Calvin se separan en el tiempo. Los estomas
pueden permanecer de esa manera cerrados durante parte del día para minimizar
la pérdida de agua. Durante la noche, con temperaturas más bajas, se abren para
tomar el CO2 y fijarlo en forma de malato, que se guarda en las vacuolas. Ya
durante el día se libera y mediante RuBisCO, la enzima clave en la fase oscura,
de manera análoga a las plantas C3 es transformado.
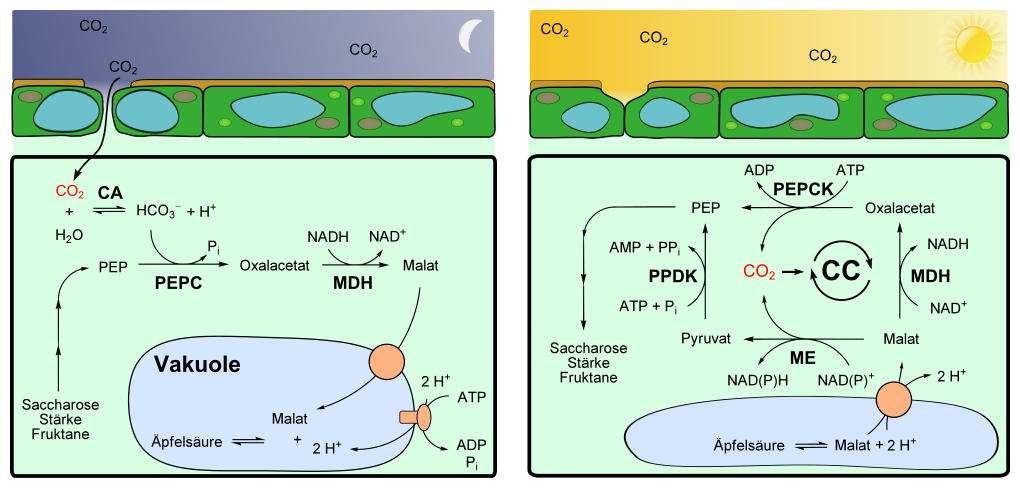 |
Esquema del metabolismo
ácido de las crasuláceas durante la noche (a la izquierda, fase I) y
durante el día (a la derecha, fase III).
CA Anhidrasa
carbónica-α.
CC Ciclo de
Calvin.
PEP Fosfoenolpiruvato.
PEPC Fosfoenolpiruvato
carboxilasa.
PEPCK Fosfoenolpiruvato
carboxiquinasa.
MDH Malato
deshidrogenasa.
ME Malato
deshidrogenasa (descarboxilante) = Malatoenzima.
PPDK Piruvato
fosfato diquinasa.
|
Fijación del
dióxido de carbono por la noche
Por la noche se logra
la fijación de CO2 en ácido fosfoenolpirúvico (PEP) mediante una
PEP-caboxilasa, parecido a como lo hacen las plantas C4. La enzima condensa
carbonato (HCO3-) con el PEP. Esto requiere la obtención de PEP a partir de
almidón, otros tipo de plantas utilizan azúcares solubles como sacarosa o
fructosano. Las plantas CAM deben tener suficiente PEP a disposición para que
una ración nocturna de dióxido de carbono pueda ser fijada. Para ello durante
el día se repone las reservas de almidón, para suministrar el suficiente PEP
por la noche.
En soluciones acuosas
se encuentra en una proporción igual el dióxido de carbono y el HCO3, pero esta
transformación es muy lenta. Una anhidrasa carbónica acelera la reacción para
equilibrar la proporción de HCO3, que posteriormente, mediante la acción de la
PEPC es transformado a oxalacetato (OA). Después, el OA formado es reducido por
la NAD-malato deshidrogenasa a L-Malato. Con ello se oxida NADH a NAD+.
El malato se
transporta a las vacuolas para ser almacenado. Las vacuolas de las plantas CAM
son mucho más grandes que la de otras especies y pueden almacenar en
comparación una alta concentración de malato (hasta 0.2 M), lo que genera una
alta capacidad de almacenamiento. Al depositar el malato en las vacuolas mantiene
los niveles de pH constantes en el citoplasma.
Para transportar el
malato en contra del flujo por el gradiente de concentración, se transportan
dos protones por cada molécula de malato bajo consumo de ATP. La reacción se
cataliza por una ATPasa de clase V que se localiza firme en el tonoplasto. Por
un transporte activo secundario, sigue el malato a los protones mediante un
transportador dicarboxilato.
El almacenaje sucede
en la forma protonada del malato, el ácido málico, que por las condiciones
ácidas se encuentra en su forma no disociada. Lo que tiene como ventaja un
valor osmótico inferior. En las vacuolas de las células de los estomas se
guarda malato de potasio, que en comparación ejerce una mayor presión osmótica
. Mientras que el pH de muchas C3 y C4 está en 5.5,10 puede alcanzar en las
plantas CAM, a causa de la concentración de ácidos, el valor de 3. Por ello por
la mañana tienen un sabor agrio.
 |
La fotosíntesis en
las plantas CAM durante el día y
la fotosíntesis en
las plantas CAM durante la noche.
|
Liberación del
dióxido de carbono durante el día
Durante el día el
proceso se invierte, el CO2 se libera. El malato almacenado es enviado fuera de
la vacuola hacia el citoplasma junto con dos protones mediante un
transportador.11 Es posible que el ácido málico con carga neutra se difunda por
la membrana directamente.12 2 Durante el día de esa manera el pH en la vacuola
vuelve a subir entre 7.5 y 8.2 A este proceso se le conoce como
desacidificación.
La descomposición del
malato y la liberación del CO2 se da como en el caso de las plantas C4 de
manera diferente de acuerdo a la especie, lo que refleja la formación de las
CAM en diferentes taxones. Los caminos existentes son: la enzima malato
deshidrogenasa dependiente de NADP (en cáctus y agaves), la enzima malato
deshidrogenasa dependiente de NAD (plantas Crassula) y la fosfoenolpiruvato
carboxiquinasa (por ejemplo en las bromelias, liláceas, asclepiadaceas).
En las plantas con la
enzima malato deshidrogenasa dependiente de NADP se descarboxila el L-malato
proveniente de la vacuola a piruvato. Esta reacción se inicia en los
cloroplastos. Con ello se reduce el NADP+ a NADPH.
Algunas plantas
tienen una enzima malato deshidrogenasa dependiente de NAD mitocondrial, que
utiliza NAD+ en lugar de NADP+ como cofactor. La reacción ocurre de manera
similar a la previamente mencionada. Pero el piruvato no se transporta de la
mitocondria al citoplasma, sino en un paso intermedio como L-alanina a los
cloroplastos. Ahí se transforma a PEP y se devuelve al citoplasma. La formación
de la alanina y su reacción inversa es catalizada por una alanina transaminasa.
Las plantas CAM con
la fosfoenolpiruvato carboxiquinasa presentan dos ciclos. Junto con el ciclo
NAD-ME también tienen una PEP carboxiquinasa (PEPCK; EC 4.1.1.49). El L-malato
es oxidado bajo consumo de NAD+ a oxalacetato por una enzima NAD-malato-DH.
Después el oxalacetato es descarboxilado directamente a PEP por la PEPCK, esto
bajo consumo de ATP.
Por las reacciones
previamente mencionadas el CO2 liberado entra en el ciclo de Calvin y es
transformado a ácido 3-fosfoglicérico, que lleva a la formación de
monosacáridos (D-glucosa, D-fructosa), seguidos por la formación de
polisacáridos (sacarosa). Por la reación de descarboxilación el piruvato
formado bajo consumo de ATP es transformado en PEP por la piruvato, fosfato
dikinasa. Por último el PEP toma el camino de la gluconeogénesis y se obtiene
almidón, sacarosa o fructanato.
Debido a que la
mayoría de los carbohidratos formados abastecen los requerimientos nocturnos
con consumo del almidón almacenado, la ganancia de las plantas CAM en la
fotosíntesis es muy poca. También el tamaño de la vacuola limita lo que se
puede almacenar de malato. Ambos factores provocan que el aumento en biomasa
esté limitado.
Por la transformación
y descomposición del malato hay un cambio en el pH de 7.5 durante el día a 3
por la noche. Debido a este cambio se habla de un ritmo ácido diurno. Este
ritmo se comprobó por primera vez en la Kalanchoe daigremontiana en 1984.
No hay comentarios:
Publicar un comentario